


 2020.07.20
2020.07.20  2956次
2956次
一、前言
分子+疾病+调控,是我们科研的惯用套路,如何在这个框架下产出精彩的故事和高分文章呢?今日我们以这篇文章为例:胰岛素样生长因子2 (IGF2)通过细胞外处理蛋白质聚集物来预防亨廷顿病【Insulinlike growth factor 2 (IGF2) protects against Huntington’s disease through the extracellular disposal of protein aggregates; 2020; IF=14.251】。
二、文章速读
受损的神经元蛋白平衡是许多神经退行性疾病的一个显著特征,突出了内质网(ER)功能的改变。之前报道过靶向转录因子XBP1是ER应激反应的关键介质,可延缓疾病的进展,减少各种神经变性模型中的蛋白质聚集。为了确定在XBP1缺失下可能解释神经保护作用的疾病修饰基因,文章对这些动物的大脑皮层和纹状体进行了基因表达检测,发现胰岛素样生长因子2 (Igf2)是主要的上调基因。文章研究了IGF2信号在亨廷顿病(HD)模型中对蛋白质聚集的影响作为证明。细胞培养研究显示,IGF2处理降低了突变体亨廷顿蛋白和聚谷氨酰胺肽的细胞内聚集物的负荷。这些结果在HD患者和脊髓小脑失调患者的诱导多能干细胞(iPSC)培养基棘神经元中得到验证。突变亨廷顿蛋白水平的降低与细胞内蛋白质半衰期的降低有关。IGF2引发的异常蛋白聚集水平下降与自噬活性和蛋白酶体途径无关,而这两种途径是清除突变体亨廷顿蛋白的主要途径。相反,IGF2信号通过外泌体和微泡促进可溶性突变亨廷顿蛋白的分泌,涉及肌动蛋白动力学的改变。用基因疗法将IGF2注入HD小鼠大脑,在三种不同的动物模型中,突变体亨廷顿蛋白的水平显著下降。此外,对HD患者死后脑组织和血液样本的分析显示IGF2水平降低。本研究发现IGF2是HD中解除管制的相关因子,作为一种抑制异常蛋白种类积累的疾病调节剂。
三、文章背景
❶ 蛋白质错误折叠和聚集是多种神经退行性疾病的先兆,如HD。分子伴侣常驻于胞液和ER以确保新合成的蛋白精确折叠。质控机制通过蛋白酶体、溶酶体、自噬途径去除错误折叠的蛋白。随着年龄的增长,蛋白酶体网络的容量减少,可能会增加蛋白聚集和神经退化。神经退行性疾病中蛋白质稳定网络改变的主要节点之一涉及ER,是细胞中蛋白质折叠和质量控制的主要部位。内质网应激引发了一种快速而协调的信号通路,即未折叠蛋白反应(UPR),以加强适应性程序并恢复蛋白稳定。内质网不可修复的应激通过细胞凋亡导致细胞死亡。
❷ UPR由激活的特殊应激转导子启动,强调肌醇需要的跨膜激酶/内切酶(IRE1α)是最保守的信号分支。IRE1亚型是一种位于ER的激酶和核糖核酸内切酶,在激活时,它控制编码X-box结合蛋白1(XBP1)的mRNA的加工,导致表达稳定和活性,转录因子称为XBP1s。XBP1s上调蛋白折叠相关基因、质控、ER易位和ERAD。在秀丽隐杆线虫中,神经元中XBP1的表达控制着机体的衰老,这可能与调节正常衰老的经典信号通路包括胰岛素样生长因子(IGF)-1和FOXO (DAF16)通路相互干扰。作者此前观察到在HD模型大脑发育过程中遗传阻断IRE1α/XBP1通路可导致神经保护,可以用减少异常蛋白聚集物积累的激元适应变化来解释。这些结果说明了XBP1和UPR在神经退行性疾病进展中的更广泛的意义,强调了确定介导神经保护的作用机制的必要性。
四、方法技术
1. XBP1基因敲除鼠
2. 实验动物大脑皮层和纹状体进行微阵列(SAM)分析
3. qRT-PCR
4. 细胞转染
5. 自噬、蛋白酶体检测
6. Western Blot
7. 脉冲追逐实验
8. mHtt分泌与点位印迹检测
9. 囊泡分离和纳米粒子跟踪分析
10.定量蛋白质组学
11.Rac1 pull-down和肌动蛋白动力学
12.腺病毒载体构建及体内外感染
13.模型小鼠旋转测试
14.组织免疫荧光
15.切片显微定量
16.HD患者血清ELISA
17.神经干细胞(NSC)诱导分化中型多脊神经元(MSN)
18.脊髓小脑共济失调3类(SCA-3)患者真皮成纤维细胞诱导多能干细胞(IPSC)生成
19.IPSC衍生神经元生成
20.神经元兴奋性刺激
21.免疫组化
五、实验结果
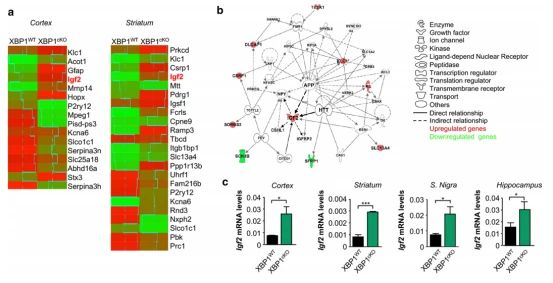
1.Igf2在XBP1敲除的小鼠脑内上调
➀ 获取XBP1敲除小鼠大脑皮层和纹状体进行基因表达分析,得到差异表达基因,在两个脑组织之间进行比较,大脑皮层和纹状体之间的基因表达变化没有重叠,只有Igf2的表达在两个脑区被改变,在XBP1cKO小鼠中表达更高。
➁ 独创性通路分析(IPA)提示Igf2与与Htt相关的神经功能障碍相关的关键生物学通路与淀粉样前体蛋白之间存在关系。第二次IPA检测发现Htt是几个靶基因(Igf2、Gfap、Mmp14、Mpeg1和Serpine3)失链且p值最低的上游调控因子的候选基因。
➂ real-time PCR验证了XBP1cKO小鼠大脑中Igf2表达水平的变化,并检测到与野生型同窝小鼠相比Igf2有显著增加。
2.IGF2减少polyQ和mHtt聚集
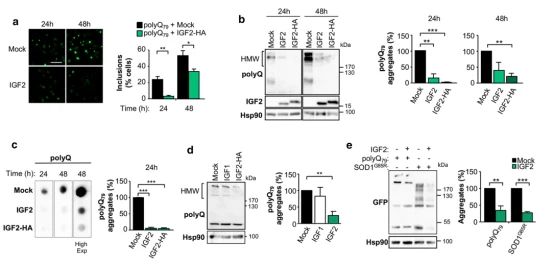
➀ 为确定IGF2表达对异常蛋白积累的影响,将polyQ79-EGFP表达载体与IGF2或空载体(Mock)一起转染Neuro2a细胞,分析细胞内包涵体。发现当与IGF2载体共转染时,polyQ79-EGFP包涵体的数量显著减少。
➁ 为了评估对聚集的影响是否针对IGF2,测量共转染IGF1表达载体的细胞中polyQ79-EGFP的积累。通过一系列的实验说明IGF2的表达降低了polyQ79聚集物和HMW物种的水平。
➂ 为了确定IGF2是否在细胞外细胞室发挥作用,用富含IGF2的条件培养基处理表达polyQ79-EGFP的细胞。还包括了一个表达突变亨廷顿蛋白跨越外显子1的片段,有85个CAG重复的构建(GFP-mHttQ85),由IGF2或空载体转染Neuro2a细胞24 h生成的IGF2富集培养基预处理的神经2a细胞中瞬时表达polyQ79-EGFP或GFP-mHttQ85。IGF2富集处理的细胞观察到明显减少蛋白质聚合和夹杂物。表明IGF2可能发挥其功能以旁分泌的方式,可能通过与膜受体结合激活信号通路。此外,用胰岛素处理细胞并没有降低polyQ79-EGFP HMW物种的水平,与IGF1过表达的结果相似。
➃ 为了评估IGF2是否可以降低预形成聚合物的含量,表达polyQ79-EGFP或GFP-mHttQ85 24小时,然后用IGF2富集或对照培养基处理Neuro2a细胞。结果发现细胞暴露于IGF2条件下的培养基可减弱错误折叠polyQ79-EGFP或GFP-mHttQ85物种的负载,这表明IGF2可能会触发异常错误折叠蛋白的降解或分解。
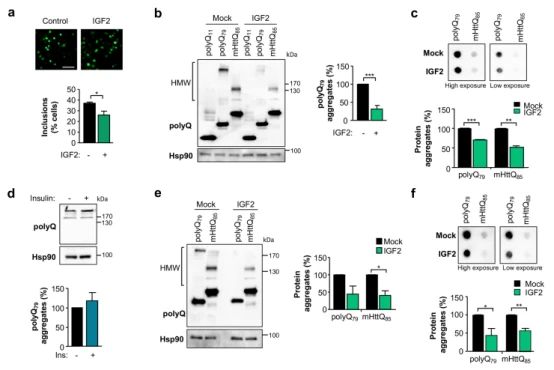
3.IGF2减少了膨胀的聚谷氨酰胺蛋白的聚集
➀ 为了确定IGF2表达对表达全长度mHtt的细胞模型的影响,转染HEK293细胞与表达载体的myc标记版本的全长mHtt重复103谷氨酰胺(FL HttQ103-myc)连同IGF2或空载体(Mock)。通过western blot分析观察到IGF2的表达导致FL HttQ103-myc的单体不溶性物种和洗涤不溶性物种的水平显著降低。用dot - blot方法分析细胞裂解液时,也证实了上述结果。总的来说,IGF2的表达导致全长mHtt的单体和HMW形式的水平下降。
➁ 为了确定了IGF2在不依赖于过度表达的人类HD模型中可能的影响,该模型使用了来自患者的诱导多能干细胞(iPSCs)。因此,使用来自HD患者和对照组的iPSC生成了人中棘神经元(MSNs)。为了表达IGF2,使用2型血清将其cDNA包装到腺相关病毒载体(AAV)中,该载体对神经元具有高度的趋向性。用AAV-IGF2或AAV-Mock转染单分散微球培养17天,western blot检测mHtt水平。值得注意的是,IGF2的表达导致了HD衍生的MSNs中mHtt总水平的下降。对照组的单分散二氧化硅微球培养物并没有改变野生型Htt的水平。
➂ 为了验证IGF2表达对其他疾病相关基因水平的可能影响,我们使用从SCA3患者获得的iPSC生成神经元培养物,并将其暴露于IGF2富集的条件细胞培养液中。细胞暴露于谷氨酸导致突变体Ataxin-3的异常聚集,在洗脱剂不溶性部分中通过突变蛋白的积累来反映。将细胞暴露在富含igf2的培养基中会导致突变体Ataxin-3聚集物的水平降低。因此,IGF2在减少含聚谷氨酰胺蛋白的异常聚集方面具有更广泛的作用。
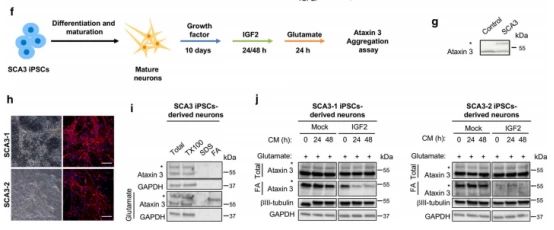
4.IGF2降低细胞内可溶性mHtt的半衰期
细胞暴露于IGF2诱导的mHtt聚集稳态水平的降低可能是合成速率的衰减或降解的增加。在HEK293T细胞中进行了脉冲标记实验,将mHttQ43-GFP构建物与IGF2或空载体(Mock)共转染。为了评估转译率,在总周期60分钟内,以S35标记的甲硫氨酸和半胱氨酸的脉冲增加间隔。这些实验表明,在表达或不表达IGF2的细胞中,mHtt的生物合成是相同的。此外,一般蛋白的合成不受IGF2表达的影响。然后,用脉冲追逐法监测细胞提取物中mHttQ43-GFP水平的衰减,以消除其半衰期。IGF2的表达显著降低了细胞提取物中mHttQ43-GFP的半衰期,但不影响蛋白质组的整体稳定性。在这些实验中,没有观察到mHttQ43的HMW物种的存在,这表明IGF2正在影响可溶性或单体形式的稳定性。总之,IGF2信号降低了总mHtt的含量,并缩短了细胞内蛋白质的半衰期。
5.自噬和蛋白酶体途径不参与IGF2诱导的polyQ聚集物的减少
➀ HEK293T细胞与mHttQ43-GFP和IGF2表达载体共转染24小时后,用放射性脉冲在抑制剂存在的情况下进行追赶实验。氯喹和硼替佐米都没有恢复IGF2对mHtt稳定性的影响。
➁ 在基础条件下,蛋白酶体抑制剂或MG132处理细胞后,细胞内包涵体数量略有增加,但在抑制剂存在的情况下,IGF2表达细胞内的polyQ79-EGFP包涵体数量没有变化通过western blot和flter trap分析,分别测量了polyQ79-EGFP HMW的种类和聚集物,从而验证了这些在Neuro2a细胞中的观察结果。在IGF2处理的细胞中,抑制蛋白酶体后polyQ HMW物种形成的基础增加消失了。这些结果表明IGF2信号可能会改变细胞内聚合/可溶形式的polyQ的比例,或者它可能会改变异常polyQ物种的方向,以介导其清除。
➂ 在神经元自噬上调的情况下,我们评估过表达IGF2的细胞的自噬激活。在有无polyQ79-EGFP表达载体的情况下,用IGF2转染Neuro2a细胞。然后用氯喹抑制溶酶体活性后监测自噬,分析LC3和p62水平。结果发现IGF2不刺激Neuro2a细胞自噬,但略有减少LC3-II的积累。同样,用氯喹处理细胞也不能恢复蛋白质。总的来说,这些结果表明,IGF2处理诱导的polyQ79-EGFP聚集量的减少与蛋白酶体和自噬/溶酶体途径无关。IGF2可能降低可溶性和单体形式的mHtt或polyQ的水平,降低它们的聚集水平,通过转移细胞内的平衡向非聚集形式。另外,IGF2信号通路可能会使异常蛋白种类的清除转向一种独立于蛋白酶体和大自噬/溶酶体活性的替代途径。

6.IGF2信号通过细胞外小泡触发polyQ分泌
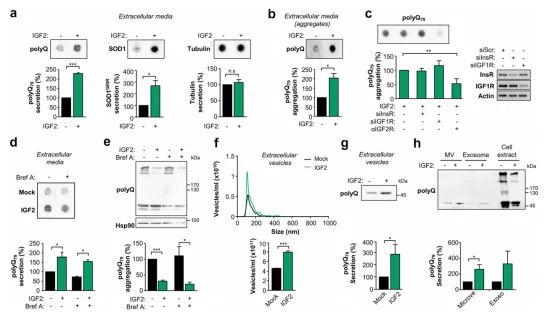
➀ 使用dot blot方法分析培养基时,IGF2表达的细胞中polyQ79-EGFP的分泌增强。对细胞培养基中SOD1G85R水平的分析显示,IGF2可诱导其分泌,而胞质标记物如微管蛋白未见变化,表明细胞未裂解或胞质泄漏。尽管信号很低,使用flter trap检测到细胞外基础水平存在大的polyQ79-EGFP聚集物,这一现象在表达IGF2的细胞中增加了两倍。
➁ 使用siRNAs来干扰胰岛素和IGF1受体(InsR和IGF1R阻断抗体靶向IGF2受体(IGF2R),拮抗IGF2R几乎降低了聚Q79-EGFP分泌水平的一半,而INSR或IGF1R的敲除则降低了一半。这些结果表明,IGF2信号通过IGF2R的参与降低了聚Q79-EGFP在细胞中的负载,导致了polyQ蛋白细胞外处理。
7. IGF2通过外显子和微囊泡刺激聚Q分泌
➀ 用BrefeldinA阻断ER至高尔基体,不能抑制IGF2刺激细胞中聚Q79-EGFP的分泌。然后探索细胞外小泡中聚Q79-EGFP的存在,使用纳米粒子跟踪分析系统来确定囊泡在细胞培养基中的大小分布和浓度表达IGF2的细胞,发现IGF2表达细胞中囊泡的释放增强,平均直径在30~150 nm之间,而一小部分(
➁ 通过顺序离心分离微囊和外胚层富集组分。在表达IGF2的细胞中,在这两个组分中检测到聚Q79-EGFP水平的增加。聚Q79-EGFP的主要单体和可溶性形式被分泌到微泡和外泌体,没有洗涤剂不溶性HMW物种。因此,IGF2信号通过外显子和微囊泡促进聚Q79-EGFP的分泌,降低细胞内聚Q79-EGFP的水平。
8.细胞骨架重构有助于IGF2诱导的polyQ分泌
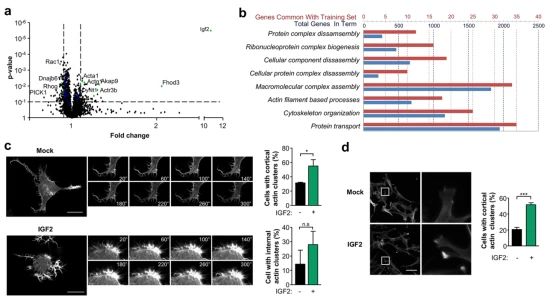
➀ 为了表征IGF2刺激下的蛋白质组重构,我们应用串联质量标记-多维蛋白识别技术(MuDP IT)定量蛋白质组学。鉴定了总共199个蛋白质,它们的表达水平发生了变化,p值0.1倍的变化。IGF2的表达不会导致蛋白质组学的改变。然而,功能富集分析表明,IGF2的表达触发了与大分子复合物相关的蛋白质的波动、拆卸、蛋白质转运和细胞骨架重组。由IGF2修饰的一组蛋白质与肌动蛋白细胞骨架动力学和调控有关,包括几个肌动蛋白 结合蛋白、RhoGTPase、波形蛋白、动力蛋白激活蛋白和动力蛋白等因素。
➁ 由于肌动蛋白细胞骨架是调节蛋白质分泌和囊泡贸易的关键,探讨IGF2是否对肌动蛋白细胞骨架的调节有任何影响,结果显示IGF2处理在几分钟内产生了非常快的肌动蛋白动力学和细胞形态的变化, 在细胞胞浆中出现肌动蛋白簇和flopodia的发育增加。这些结果是通过可视化肌动蛋白细胞骨架在小鼠胚胎胚胎纤维细胞中融合的。
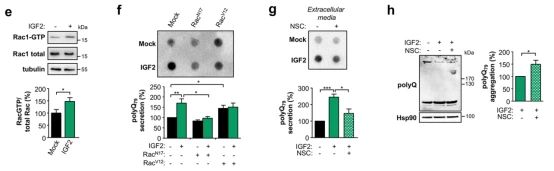
➂ 考虑到RAC1在细胞骨架动力学中的中心作用,评估了IGF2刺激下的RAC活性。结果表明,与GTP偶联的活性Rac1的量迅速增加,治疗IGF2。
➃ 测试肌动蛋白动力学对聚Q79-EGFP肽释放到细胞外空间的功能贡献。神经2a细胞瞬时转染两个显性阴性 (Rac117N)和Rac1的本构活性(Rac1V12)形式。Rac117N能够阻断IGF2增强的聚Q79-EGFP分泌。相反,Rac1V12的表达促进了在基础水平上分泌OlyQ79-EGFP,而在IGF2刺激的细胞中没有观察到差异,提示该途径已经完全激活。用NSC23766抑制RAC1-GTPase活性,显著降低聚Q79-EGFP的分泌。这些效应与在NSC23766存在下用IGF2处理的细胞内聚Q79-EGFP的细胞内HMW物种水平的增加平行。这些结果提示IGF2信号触发细胞骨架动力学的快速变化,导致聚Q79-EGFP肽进入细胞外空间。
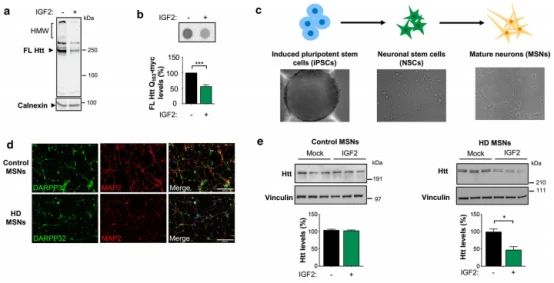
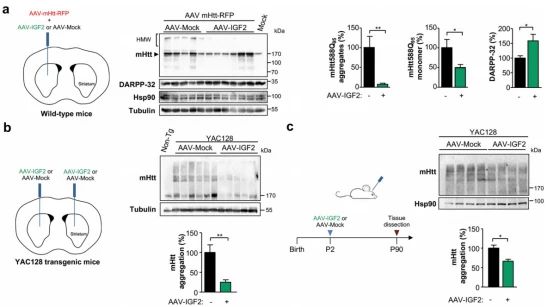
9.体内IGF2减少MHtt聚集
➀ 为了确定IGF2在体内对mHtt聚集水平的可能影响,进行了单侧立体定向注射AAVS混合物,以交付Htt588Q95-RFP与IGF2或空矢量(Mock)进入纹状体。在AAV分娩后两周,对小鼠进行安乐死,并解剖纹状体进行生化分析。在成年小鼠中,IGF2的局部表达导致mHtt聚集明显减少。与细胞培养实验一致,注射AAV-IGF2的HD小鼠的大脑中也减少了mHtt的单体形式。为了评估过度表达IGF2对神经元存活的影响,测量了DARPP-32水平,作为中等刺状神经元活力的标志。与对照病毒相比AAV-IGF2处理的动物中检测到DARPP-32明显更高的水平。
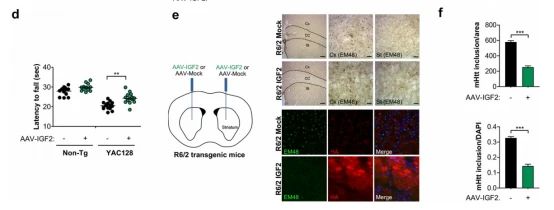
➁ 考虑到使用病毒HD模型获得的阳性结果,随后评估了基因治疗策略,在杂合状态下在YAC128转基因模型中提供IGF2。进行了双侧立体定向注射将AAV-IGF2或空载体导入3个月大的YAC128小鼠纹状体,然后在四周后对脑提取物进行生化分析,发现纹状体中全长mHtt表达减少,平均下降近80%。注射出生后第1天或第2天(P1-P2)YAC128小鼠的DAV-IGF2或对照载体,并在3个月后解剖纹状体。免疫印迹分析表明IGF2的表达显著降低了YAC128动物大脑中mHtt的总水平。最后,确定了使用基于IGF2的基因治疗对实验性HD临床进展的功能后果,结果发现随着时间的推移,IGF2进入神经系统改善了HD转基因小鼠的平均运动性能。
➂ 为了补充我们的体内验证,用第三个模型,R6/2小鼠,一个转基因HD模型,表达了人类Huntingtin的第1外显子,含有~150个CAG重复,这是ALLO 胞内m Htt包裹体的可视化。评估了IGF2在R6/2小鼠大脑中的作用,在AAVIGF2给药时,观察到mHtt阳性夹杂物含量的强烈降低, 这些实验的量化表明,用AAV-IGF2处理的R6/2小鼠的大脑中有近60%的显著减少。综上所述,这些结果表明,人工执行IGF2在大脑中的表达减少了不同HD模型中异常的蛋白质聚集。
10. HD患者纹状体和血样中IGF2水平降低
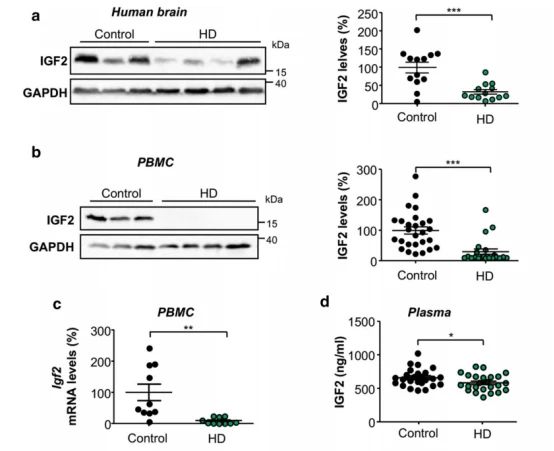
➀ 进一步探讨HD患者标本中IGF2表达的可能改变。Western blot分析结果显示与对照组相比,显示HD患者大脑中平均IGF2水平降低了近66%。
➁ 考虑到HD死后脑组织中IGF2水平的意外下降,评估了HD患者外周血单个核细胞(PBMCs)中IGF2的存在。结果发现HD衍生细胞IGF2水平降低,用western blot分析显示其蛋白水平下降了近80%;而其mRNA水平下降了近90%。综上所述,这些结果表明HD患者的脑和血细胞中IGF2水平急剧降低。

六、思路总结
✦ 选定一个分子X
✦ 确定其作用的疾病A(潜在的治疗作用)
✦ 选择X可能通过机制M作用于A
✦ 动物模型证明X表达变化(动物疾病模型、基因敲除模型)
✦ 体外模型论证X的M机制(选择M可量化及可观察的指标)
✦ A样本中X水平与对照组不同
科研套路深入分析及实验方案设计,
广州辉园苑为您保驾护航,
让思路更清晰、科研更顺利!
一站式整体科研服务平台
做最真实的数据,提供最真诚的服务
如有需要,欢迎联系我们:
020-29039963/qq:2951428216
 返回列表
返回列表