


 2020.07.13
2020.07.13  5282次
5282次
转录因子(Transcriptionfactor,TF)是一种能与特异DNA序列结合的蛋白,可以单独或与其他蛋白形成复合体,提高或阻断特定基因对RNA聚合酶的招募,调控基因的表达。转录因子的特点是它包含一个或多个DNA结合域(DNA-binding domain,DBDs),通过这些结合域与基因附近的DNA序列结合,从而完成调控。下面结合文章介绍转录因子的研究方法。
01、案例文章简介
案例文章:
Ahr-Foxp3-RORt axis controls gut homing of CD4+T cells by regulating GPR15. Sci Immunol. 2020, 5(48). (IF=13.44)

GPR15是一种趋化因子受体,在T细胞向大肠归巢的过程中发挥重要作用。这项研究发现小鼠大肠Treg中的GPR15表达水平最高,芳香烃受体(Ahr)可通过增加GPR15的表达促进Treg向大肠的归巢,而Foxp3及RORγt在Ahr介导的GPR15表达上调中分别起到正向及负向调控作用。另外,在人结肠Treg中也可重复上述发现。该研究提示,Ahr-Foxp3-RORγt轴可通过调控GPR15的表达调节Treg的肠道归巢,从而影响肠道免疫稳态。
02、案例文章的研究内容
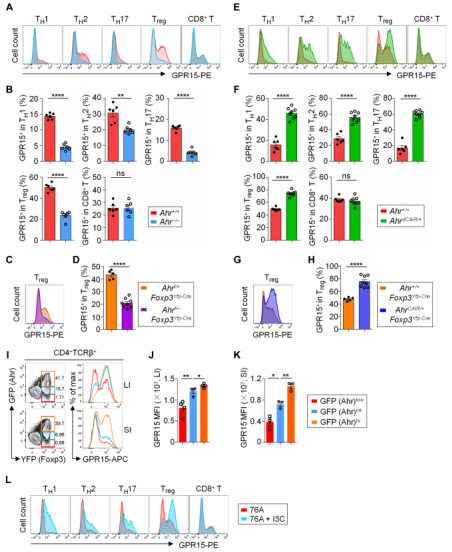
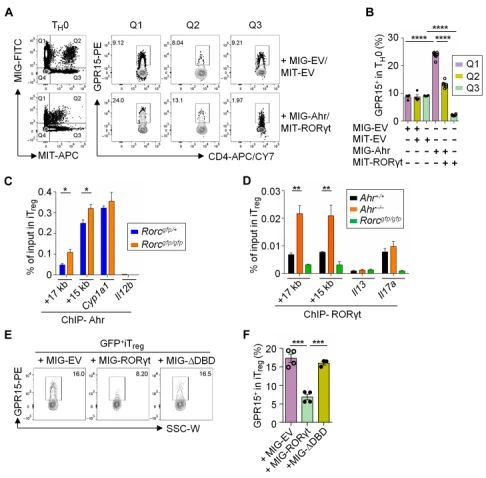
GPR15在小鼠大肠Treg中表达量最高,并且受Ahr控制。GPR15是一种孤儿鸟嘌呤核苷酸结合蛋白(G蛋白)偶联趋化剂受体(GPCR),最初是基于其与其它GPCR家族成员的相似性而被发现的,也被称为HIV或猿猴免疫缺陷病毒共受体。最近有研究表明,GPR15对小鼠和人的调节性T细胞(Treg)和效应T细胞(Teff)的肠道归巢至关重要。芳香烃受体(Ahr)是一种环境传感器,可检测外源性配体,如环境毒素(如二恶英),以及宿主细胞、微生物群和饮食(如色氨酸代谢物)产生的内源性配体。通过流式细胞术检测GPR15蛋白在各种T细胞中的表达水平,发现Treg中GPR15的表达水平最高。在Ahr缺陷小鼠的CD4+ T细胞中,GPR15的表达水平降低。而在在Ahr过量表达小鼠的CD4+ T细胞中,GPR15的表达水平升高。
Ahr通过直接与Gpr15基因座结合调节GPR15表达。利用ChIP-seq在iTreg 和TH17细胞中发现Gpr15基因是Ahr最重要的全基因组靶点之一。Ahr在Gpr15基因座具有两个实质性的结合峰(在距转录起始点+15和+17 kb处)。
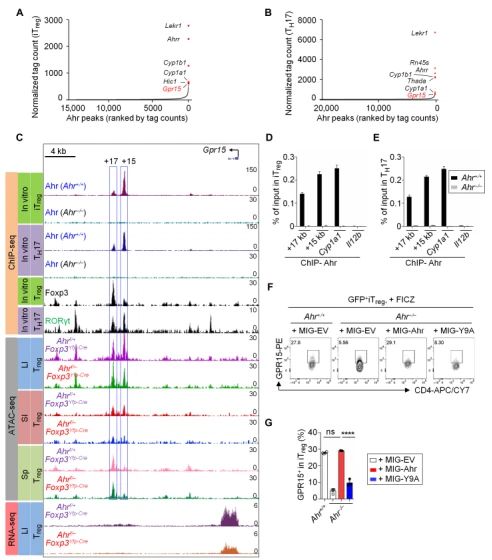
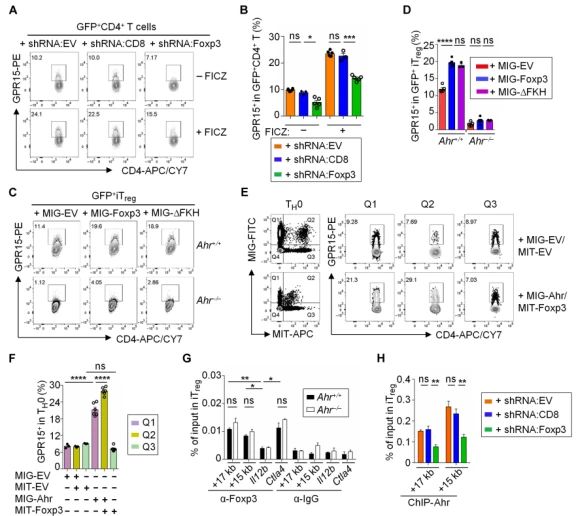
Foxp3与Ahr协同作用,正调节GPR15的表达。在CD4+ T 细胞中用shRNA干扰Foxp3,GPR15的表达水平降低。在来自Ahr+/+小鼠的iTregs细胞中强制表达Foxp3可显著提高其GPR15的表达,在来自Ahr-/-(Ahr缺陷)小鼠的iTregs细胞中却没有这种效果。在TH0条件下培养的分选CD4+T细胞中同时表达Ahr和WT Foxp3或Foxp3△FKH可以增加GPR15的表达,而Foxp3单独表达并不影响GPR15的表达。ChIP-qPCR表明Foxp3在Gpr15基因座的结合位点与Ahr相同(距转录起始点+15和+17 kb处)。Ahr缺陷不影响Foxp3在Gpr15位点的结合,但是在iTreg中敲低Foxp3后,Ahr向Gpr15基因座的招募减少。
RORγt负向调节GPR15表达。在RORγt缺陷Treg中,GPR15的表达增加。RORγt和GPR15表达的负相关系在Ahr充足和Ahr缺陷Treg中都很明显。此外,与来自RORγt充足的CD4+ T细胞分化的iTregs和TH17细胞相比,来自RORγt缺陷的CD4+ T细胞具有更高的GPR15表达。
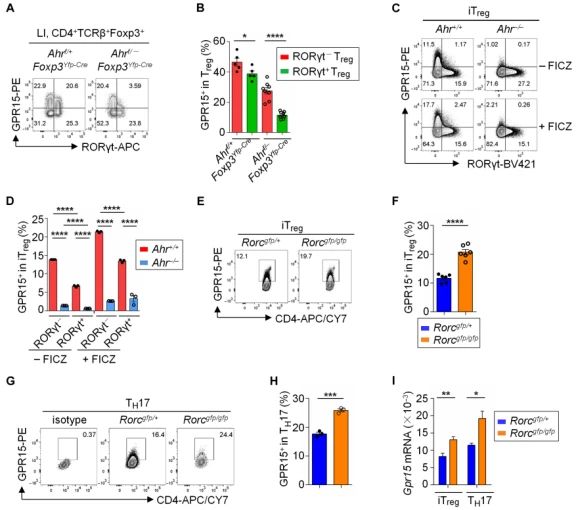
RORγt拮抗Ahr在Gpr15基因座的DNA结合。在从RORγt缺陷小鼠分化的iTregs或TH17细胞中,Gpr15基因座的Ahr募集显著增强,而在没有Ahr的情况下,Gpr15基因座的RORγt结合显著增加,提示Ahr和RORγt与Gpr15基因座结合存在竞争。此外在RORγt缺陷iTregs中表达RORγt抑制GPR15表达。
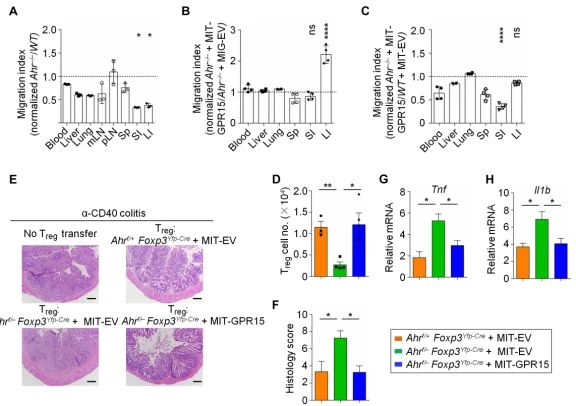
Ahr通过调节GPR15促进Tregs向肠道归巢。在大肠(LI)和小肠(SI)中,归巢的Ahr缺陷iTreg相对于WT对照组减少了大约三倍。GPR15的强制表达显著增强了Ahr缺陷Treg向LI的归巢,但不增强到包括SI在内的其他器官的归巢,这与GPR15在Treg大肠归巢中的关键作用一致。此外, 在LI中WT-iTreg和GPR15恢复表达的Ahr缺陷iTreg的归巢能力相当(而不是在SI中),这表明GPR15在Ahr缺陷iTregs中的表达受损是其向LI归巢缺陷主要机制。在Ahr缺陷的iTregs中强制表达GPR15促进其在炎症期间归巢至LI。此外,强制表达GPR15的Ahr缺陷iTreg的过继性转移显著抑制了肠道炎症,表现为减轻了肠道组织学变化和降低了促炎细胞因子(即Tnf和Il1b)的表达。
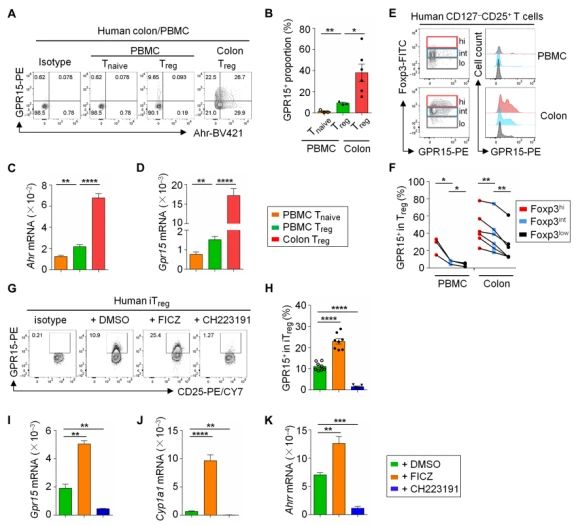
Ahr信号促进GPR15在人Treg中表达。与之前的报告显示GPR15在人类Treg中的低表达不同,本研究检测到GPR15在所有患者样本中的表达。与外周血单个核细胞(PBMCs)相比,结肠Treg的 GPR15表达显著增高。人Treg中Ahr mRNA表达的增加与Gpr15表达的增加以及其他Ahr靶基因(Cyp1a1和Ahrr)的表达增加有关,这与Ahr促进Gpr15表达的作用相一致。此外,通过蛋白质染色分析GPR15和Foxp3的表达,发现Foxp3的表达与人CD127-CD25+ T细胞中GPR15的表达呈正相关。加入Ahr的配体FICZ显著增强了iTreg中GPR15和其他Ahr靶基因的表达,但不增强Ahr的表达,而用一种特异性Ahr拮抗剂CH223191治疗可消除人iTregs中GPR15的表达。
03、转录因子研究的技术手段
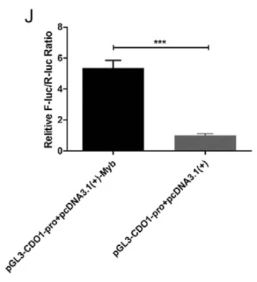
❶ 双萤光素酶实验
某些转录因子仅与其靶启动子中的特异序列结合,这些特异性的序列被称为顺式作用元件,转录因子的DNA结合域和顺式作用元件实现非共价结合,从而对基因的表达起增强或抑制的作用。双萤光素酶报告实验(dual luciferase assay)是检测转录因子和靶启动子中的特异顺序结合的重要手段,通过检测转录因子表达水平的改变对荧光素酶基因表达水平的影响,分析转录因子能否作用于启动子。
其原理简述如下:
(1)构建一个将靶启动子的特定片段插入到萤光素酶编码序列上游的报告基因质粒,如pGL3-basic等。
(2) 将要检测的转录因子的表达质粒、干扰质粒或者siRNA与报告基因质粒共转染293细胞或其它相关的细胞系。通常会用另一种萤光素酶作为内参,避免转染效率对实验结果的影响。
(3) 加入特定的萤光素酶底物,萤光素酶与底物反应,发光,通过检测发光强度可以测定萤光素酶的活性,从而判断转录因子是否能与靶启动子片段有作用。
❷ 染色质免疫共沉淀
染色质免疫沉淀技术 (chromatin immunoprecipitation assay,ChIP)被广泛用于研究体内转录因子与靶基因启动子上特异性核苷酸序列的结合,并已成为研究染色质水平基因表达调控的最有效的方法。ChIP的基本原理是在活细胞状态下固定蛋白-DNA复合物,并将其随机切断为一定长度的染色质小片段,然后通过免疫学方法沉淀此复合体,特异性地富集目的蛋白结合的 DNA 片段。通过对目的片断的纯化与检测,从而获得蛋白质与DNA相互作用的信息。通过qPCR或二代测序,筛选与目的蛋白互作的DNA信息。ChIP-PCR用于鉴定转录因子结合到靶基因的启动子上,而ChIP-seq用于筛选转录因子直接结合的靶基因,探究转录因子的作用元件。

❸ 电泳迁移率改变试验
电泳迁移率改变试验 (electrophoretic mobility shift assay,EMSA ) 也称凝胶阻滞试验,DNA 在凝胶中的迁移率与相对分子质量及构型有关。裸露DNA与含有结合蛋白的DNA- 蛋白质复合物电泳时,裸露 DNA 迁移率主要取决于DNA本身,DNA-蛋白质复合物由于体积较大而滞留在较后位置。电泳迁移率改变试验用于体外分析转录因子与靶基因启动子的结合。将一系列序列不同的DNA片段与转录因子进行电泳迁移率改变试验,可以探究转录因子的体外结合序列。

 返回列表
返回列表